- Las blástulas se convierten en gástrulas cuando sufren el proceso de gastrulación
- el proceso de gastrulación implica un crecimiento embrionario, aumentando el tamaño. También hay una reorganización celular que lleva a la aparición de las capas germinales. Ahora aparecen dos de estas capas, el endodermo y el ectodermo
- los primeros movimientos celulares de la gastrulación son muy parecidos en todos los animales pero los mecanismos de gastrulación dependen mucho de la cantidad y disposición de vitelo
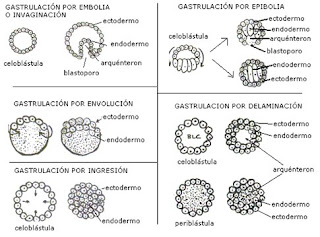
- hay varios tipos de gastrulación:
- G. por embolia o invaginación
- la sufren las celoblástulas con blastocele central
- las células del polo vegetativo se pliegan hacia dentro y se introducen hacia el blastocele mediante un proceso llamado invaginación
- de esta forma se crea una cavidad rodeada por ectodermo que se llama ARQUÉNTERON y será el futuro tubo digestivo
- el poro de entrada al arquénteron se llama BLASTOPORO y dependiendo del posterior desarrollo el blastoporó dará lugar al ano, la boca o se cerrará
- se forman dos capas germinales, la más externa es el ectodermo (que dará lugar al epitelio de la superficie corporal y al sistema nervioso) y la más externa, que constituye el arquénteron, es el endodermo (que formará el epitelio del tubo digestivo). Las evaginaciones del arquénteron forman el mesodermo (tercera capa embrionaria que formará el sistema muscular y reproductor)
- G. por epibolia
- ocurre en celoblástulas con blastocele excéntrico (desplazadas hacia el polo animal)
- no se produce invaginación porque los macrómeros del polo vegetativo no tienen movilidad
- se mueven los macrómeros del polo animal que se multiplican por mitosis y se desplazan envolviendo los macrómeros. Puede dar dos resultados:
- los micrómeros no llegan a unirse entre sí en el polo vegetativo. En esta gástrula la capa de células exterior (micrómeros) serán el ectodermo y las células internas (macrómeros) serán el endodermo. Tienen un pequeño arquénteron y un blastoporo pero desaparece el blastocele
- los micrómeros se unen en el polo vegetativo, las capas germinales son las mismas pero no hay ni blastocele ni arquénteron. El animás tendrá tubo digestivo completo pero se formará en etapas más tardías
- G. por delaminación
- ocurre en dos tipos de blástulas, en celoblástula con blastocele central y en periblástulas
- consiste en que la capa externa de células de la blástula se duplica y forma otra interna
- la capa externa será el ectodermo y la interna el endodermo y queda una cavidad, el arquénteron pero no hay blastoporo
- si sucede en una periblástula sucede lo mismo pero el arquénteron está lleno de vitelo y no hay blastoporo
- G. por ingresión
- en la celoblástula las células comienzan a multiplicarse hasta rellenar el blastocele
- la capa externa será el ectodermo y la interna el endodermo
- no hay blastoporo, arquénteron ni blastocele
- G. por involución
- el disco de células del polo animal se multiplica y forma otra capa hacia dentro
- el embrión tiene ectodermo y endodermo pero nada más.

Las capas germinativas
- en la gástrula aparece una tercera capa germinal entre el ectodermo y el mesodermo
- si la tercera capa se forma a partir del ectodermo se le llama ECTOMESODERMO O MESÉNQUIMA
- si se forma a partir del endodermo se le llama ENDOMESODERMO O MESODERMO VERDADERO
- a los animales que solo poseen dos capas germinativas se las llama DIBLÁSTICOS y a los que tienen tres TRIBLÁSTICOS. Los diblásticos no pueden generar nunca el celoma y los triblásticos pueden o no general el celoma (cavidad rodeada por tejido mesodérmico)
- mecanismos de formación del mesodermo
- esquizocelia: el mesodermo se forma a partir de una célula del endodermo próxima al blastoporo (el blastómero 4d) Esta célula empieza a dividirse por mitosis y forma dos masas macizas de células que quedan al principio flotando en el blastocele
- enterocelia: las células de la pared de arquénteron comienzan a proliferar formando dos evaginaciones hacia el blastocele (vesículas celomáticas)
- en el momento en el que ya se tienen las capas germinales éstas empiezan a dividirse para formar los tejidos y órganos
- destino de las capas germinales:
- ectodermo:
- cubierta exterior del cuerpo, el tegumento y otras estructuras derivadas de él, pelo, uñas, glándulas epiteliares. revestimiento de la boca, esmalte dental, oído interno, epitelio nasal y olfativo
- tubo neural: encéfalo, médula espinal, nervios motores
- cresta neural: ganglios sensoriales y nervios, médula adremal, ganglios simpáticos, cráneo, arcos branquiales
- extremos anterior y posterior del cuerpo que reciben el nombre de ESTOMODEO y PROTODEO
- endodermo:
- glándulas anejas al tubo digestivo
- epitelio del tracto respiratorio
- faringe, tiroides, hígado, páncreas
- tubo digestivo primitivo
- mesodermo:
- la mayoría de los órganos internos
- revestimiento de las cavidades torácica y abdominal
- órganos del sistema urogenital, uréter, riñón, gónadas, conductos reproductores
- sistema circulatorio, sangre, médula ósea, tejido linfático, músculo esquelético, hueso y cartílago del esqueleto, dermis y tejido conjuntivo
- los animales diblásticos forman estas estructuras a partir del ectodermo.

Tipos de cavidades corporales en los animales
- destino del blastocele o cavidad primaria
- el blastocele desaparece porque se rellena totalmente de mesénquima. El embrión será diblástico
- el blastocele desaparece pero no se rellena de mesodermo. Son animales triblástico acelomados, porque no presentan ningún tipo de cavidad
- el blastocele no desaparece pero una ver formado el mesodermo queda únicamente tapizando la superficie externa del animal. Al blastocele se le denomina pseudoceloma. En esta cavidad faltan los órganos. Son animales Triblásticos Pseudocelomados
- el blastocele desaparece porque aparece una segunda cavidad embrionaria, el celoma. Son animales triblásticos celomados
- el celoma es la cavidad secundaria del embrión y se forma en el mesodermo. Se forma mediante dos mecanismos:
- esquizocelia: las masas de mesodermo se ahuecan y aparece una cavidad en el interior de tal forma que la pared de cada una es el mesodermo. Las dos se van haciendo grandes hasta que entran en contacto
- enterocelia: las dos evaginaciones que salen del arquénteron se independizan, el exterior será el mesodermo y el interior el celoma. Se llega al mismo resultado que en el anterior
- no es posible saber si un animal forma el celoma por esquizocelia o enterocelia, pero en el primer caso el animal puede ser acelomado porque este tipo de formación no implica que se forme celoma, al contrario que la enterocelia que cuando se forman las evaginaciones se forma el celoma necesariamente
- diferencias entre el celoma y arquénteron
- el celoma aparece posteriormente al desarrollo que el blastocele
- una cavidad celómica está rodeada completamente por mesodermo
- el celoma siempre se dispone en cavidades laterales (como mínimo un par de ellas), lo que conduce a que los animales tengan un mesenterio dorsal y otro vental
- una cavidad celómica siempre está tapizada por peritoneo
- en un animal celomado los órganos son siempre retroperitoniales
- diferencias con un pseudoceloma
- la cavidad del animal está dispuesta radialmente en torno al tubo digestivo
- no existen mesenterios al ser una cavidad única
- cavidad tapizada externamente por mesodermo pero internamente entra en contacto con el endodermo
- los órganos internos están libres en la cavidad, flotan en el pseudoceloma
- no tienen peritoneo los animales pseudocelomados
- funciones de la cavidad corporal
- el fluido interno de la cavidad corporal actúa como un sistema de distribución, fundamentalmente de gases respiratorios y de nutrientes de todos los tejidos. Si el animal posee una pared del cuerpo fina, permeable al O2, puede atravesar la pared y llegar a la cavidad, de igual forma los nutrientes que entran por el aparato digestivo son absorbidos, se vierten a la cavidad y así se distribuyen fácilmente por los tejidos del cuero
- permite que los animales alcancen mayor tamaño que los que no la poseen. Los que no tienen cavidad tienen que tener tamaños en los que la difusión sea el mecanismo suficiente como para transportar alimentos y oxígeno
- permite una reducción de los órganos múltiples que poseen los organismos acelomados, en los cuales tiene que haber varios pares de estructuras para realizar una función (para que esa función llegue a todos los tejidos)
- permite crecimiento independiente de los órganos. Que uno de los órganos creza más que el resto dentro de la cavidad no es importante porque lo único que pasa es que aumenta la presión dentro de la cavidad
- permite el movimiento independiente de los órganos. Por ejemplo en las lombrices de tierra mientras se desplazan hacia delante el tubo digestivo lo hace en sentido contrario para expulsar las heces: la pared del cuerpo y el tubo digestivo se mueven en distintas direcciones y para ello tiene que existir un fluido entre ambos que amortigüe.
- permite una mayor coordinación. Al tener un fluido como el sistema circulatorio los productos importantes como las hormonas pueden llegar a todo el cuerpo rápidamente y simultáneamente. Esto es importante en fases de reproducción, crecimiento, metamorfosis, etc.
- todas las cavidades actúan como esqueleto hidrostático
- permiten la posesión de estructuras ereversibles, estructuras que pueden retraerse dentro del cuerpo o evaginarse. Se llaman trompas o probóscides y permiten diversificar los hábitos alimenticios.

Tipos de tejidos animales
Tejido epitelial
- el epitelio es una capa celular que tapiza una superficie externa o interna. Los epitelios tapizan todos los órganos, así como canales y conductos por los que se transportan diversos materiales y secreciones. En muchas superficies las células se modifican para formar glándulas productoras de moco, hormonas o enzimas. Los vasos sanguíneos no penetran en el epitelio por lo que éstos dependen de la difusión para alimentarse
- puede generarse a partir de cualquiera de las capas embrionarias ya que su función principal es la de protección
- como ejemplos:
- epidermis, a partir del ectodermo
- epitelio intestinal, a partir del endodermo
- epitelio pleural, a partir del mesodermo
- hay epitelios glándula es y sensoriales
Tejido mesenquimático
- todos proceden del mesodermo
- ejemplos:
- tejido óseo o cartilaginoso
- tejido adiposo
- tejido vascular (células linfáticas y sanguíneas)
- tejido conjuntivo (tejido de relleno)
Tejido muscular
- procede del mesodermo
- hay tres tipos
- muscular liso, contracción involuntaria, se encuentran fundamentalmente en las vísceras
- músculo estriado, control voluntario, lengua, piernas, etc
- músculo cardiaco, contracción involuntaria
Tejido nervioso
- procede del ectodermo
- la unidad funcional es la neurona
- la finalización de la etapa embrionaria tiene lugar por
- eclosión del huevo si es un animal ovíparo
- abandono del cuerpo materno si es vivíparo
- el individuo que nace puede tener dos morfologías
- juvenil: individuo muy similar al adulto pero de menor tamaño y que todavía no alcanza la madurez sexual. Para convertirse en adulto sufre un desarrollo directo, crece y adquiere la madurez sexual
- larva: individuo morfológicamente distinto al adulto y también carece de madurez sexual. Para alcanzar el estado adulto sufre una metamorfosis. En la mayoría de los casos de desarrollo indirecto la larva ocupa recursos y hábitats muy diferentes a los del adulto. Esto es importante porque los animales que tienen fases larvarias las utilizan como mecanismos de dispersión. Como ejemplo la mayoría de los animales marinos poseen fases larvarias y la larva es planctotrófica librenadadora, mientras que el adulto suele ser o bien sésil filtrador o bien no sésil pero bentónico carnívoro, como en la mayoría de los equinodermos. Otros ejemplos son las mariposas con adultos fluidófago y larva fitófaga.
- la cantidad y distribución del vitelo es una adaptación evolutiva. El vitelo es simplemente una adaptación que permite al embrión desarrollarse sin una fuente de alimentación externa. Los embriones con poco vitelo pueden formar rápidament un estado larvario que se alimenta por sí solo o desarrollar una placenta para la nutrición.
- la filogenia establece relaciones filogenéticas de un taxón
- la ontogenia estudia el desarrollo estudia el desarrollo de un individuo desde que nace hasta que muere
- Principio de recapitulación:
- A mediados del siglo XIX Haeckel enuncia el PRINCIPIO DE RECAPITULACION (o ley biogenética): la ontogenia recapitula (repite) la filogenia. Esto quiere decir que las etapas de desarrollo de un individuo repiten las formas adultas de todos sus antecesores. Además afirma que las etapas embriológicas en todos los animales eran las las mismas: zigoto, blástula, gástrula y que cada una de estas etapas correspondía con cada una de las fases adultas de los antecesores (zigoto a protozoo, blástula a protozoo colonial y gástrula a cnidario)
- en el desarrollo de los vertebrados pasamos por una fase en la que tenemos perforada la faringe (hendiduras faríngeas). Según Haeckel pasamos en nuestro desarrollo por una etapa de pez.
- si esto fuera así, reproducir nuestra historia evolutiva sería muy fácil, pero hay dos razones por las que ésto no es así:
- la evolución también afecta al desarrollo embrionario, en todo caso se puede decir que la filogenia de un taxón puede reflejar una sucesión de ontogenias ancestrales
- los genes de un grupo de individuos no cambian por necesidades de un individuo y eso significa que acumulamos muchos genes heredados que no son útiles (lastre filogenético)
- al estudiar la ontogenia ayuda mucho establecer la filogenia, pero no únicamente eso
- En el siglo XIX K.E. von Baer da una explicación basada en que las características tempranas del desarrollo eran compartidas de forma más amplia entre diferentes grupos de animales que los caracteres más tardíos. Los adultos de animales con ontogenias relativamente cortas o simples a menudo se parecen a estados preadultos de otros animales cuya ontogenia es más compleja, pero los embriones de los descendientes no tienen por qué parecerse a los adultos de los antecesores
Heterocronías
- cuando se compara la ontogenia de dos especies filogenéticamente próximas puede ocurrir que un determinado carácter aparezca en una de las especies en una etapa del desarrollo mucho más temprana. A este desfase se le llama HETEROCRONIA
- hay un tipo especial de heterocronía,
- el ajolote tiene branquias por dos causas:
- individuo adulto que ha conservado caracteres larvarios (a esto se le llama neotenia)
- puede ser una larva que ha adquirido madurez sexual (esto se llama paedogénesis)
- los procesos de paedogénesis pueden dar lugar a especies nuevas. Hay bastantes pruebas a favor de que en la evolución de los cordados la larva de las Ascidias adquirió madurz sexual por paedogénesis y sin sufrir metamorfosis dio lugar a un individuo adulto. Esto está a favor de que los vertebrados venimos por evolución mediante paedogénesis.
Gastrulación
{kind=link}

Gastrulación de un animal diblástico: La formación de capas germinales desde una (1) blástula a una (2) gástrula. Parte de las células del ectodermo (naranja) se movilizan hacia el interior formando el endodermo (rojo).
La gastrulación es una etapa del desarrollo embrionario, que ocurre después de la formación de la blástula, esto es, que sigue a la de segmentación o clivaje, y tiene por objeto la formación de las capas fundamentales del embrión (capas germinales):
- Ectodermo: la capa más externa de células que rodea al embrión.
- Mesodermo: células que forman la parte superior de la capa que creció hacia el interior en la blástula.
- Endodermo: capa de células más interna.
En los vertebrados a la gastrulación le sigue la fase de neurulación.
Mitosis y migración celular
Una vez formada la blástula, se produce un desplazamiento de células superficiales hacia el blastocele.Esto genera una invaginación y consecuente disminución del tamaño de la cavidad blastocélica. Esto se logra mediante el ingreso de células (a las que inicialmente se podría atribuir características ectodérmicas) por el blastoporo, en un movimiento parecido al de dar la vuelta a un calcetín. El punto de entrada de estas células conforma una abertura denominada blastoporo. Al mismo tiempo que la cavidad blastocélica disminuye, surge una nueva cavidad llamada arquénteron o gastrocele, que más tarde se convertirá en el intestino. La actividad mitótica, muy intensa a lo largo de la segmentación, disminuye aunque sin cesar nunca por completo.
Los blastómeros, o agrupaciones de ellos, emprenden migraciones considerables de las que se origina la segregación celular en dos tipos, uno de los cuales cubrirá al otro. La capa externa o ectoblasto (ectodermo), cubre la capa interna o endoblasto (endodermo). Pero la gástrula no es germen diblástico más que en los poríferos y celenterados (cnidarios y ctenóforos); en todos los demás metazoos, una capa media o mesoblasto (mesodermo) queda intercalada entre las dos capas antes mencionadas.
Así, en la blástula una parte de los blastómeros comienza a invaginarse, formándose el blastoporo. El lugar donde se produce esto, está regulado genéticamente. La invaginación progresa, e invade todo el territorio del blastocele que se va viendo reducido proporcionalmente al aumento del arquénteron o nueva cavidad que se va formando, que tiene la particularidad de estar en contacto con el exterior a través del blastoporo.
En esta etapa, el embrión se denomina gástrula y dará origen a las capas del embrión descritas anteriormente. A través del proceso de gastrulación se han formado dos capas de blastómeros, una en contacto con el exterior o ectodermo y otra en contacto con el arquénteron o endodermo y entre las dos el blastocele con el líquido blastocélico.
El proceso de gastrulación ocurre de modo diferente según el tipo de huevo y su subsiguiente segmentación. Los principales tipos de gastrulación son:
Gastrulación por invaginación o embolia
La mayoría de los filos animales presentan segmentación holoblástica y, en este caso, la blástula tiene aspecto de una bola hueca (celoblástula); la cavidad que delimita se denomina blastocele. Una blástula de este tipo experimenta la gastrulación por un proceso de invaginación, és decir, que una parte de las células ectodérmicas se invagina hacia el blastocele para formar la segunda hoja embrionaria (endodermo) y delimitar una nueva cavidad, el arquénteron que comunica con el exterior por el blastoporo. Es el tipo que se ha detallado en el apartado "Secuencia".
Ocurre cuando el huevo presenta una cantidad moderada de vitelo situado en el polo vegetativo, y la segmentación produce grandes macrómeros vitelinos. En este caso, el blastocele es más o menos virtual (estereoblástula) y la invaginación se hace dificultosa. Por este motivo, los micrómeros situados en el polo animal se dividen, proliferan, se hunden y rodean los macrómeros, hasta formarse el blastoporo en el polo vegetativo.
Gastrulación por involución Cuando la cantidad de vitelo del huevo es tan grande que las segmentación es de tipo meroblástico se forma un pequeño casquete de células o discoblástula y entonces la gastrulación se produce generalmente por involución. Este proceso consiste en que las células de la periferia del disco se hunden y se dirigen hacia atrás, bajo la capa superficial y forman un doble estrato. Es el caso típico de los cefalópodos.
Es un tipo poco frecuente. Ocurre también cuando se forma una estereoblástula. El endodermo se forma a partir de la división de las célula ectodérmicas, y de la migración y hundimiento de las mismas; la gastrulación se completa con separación de dos capas de células, una externa y otra interna, y no hay blastoporo, sino que la cavidad del arquénteron se abrirá por un proceso secundario.
Es similar al caso anterior, pero el endodermo se forma por migración de células ectodérmicas en vez de por división y migración de las mismas. En este caso, tampoco se forma el blastoporo durante el proceso. Este tipo lo presentan los cnidarios, ctenóforos y esponjas.
Distinción según número de capas
Como se ha comentado antes, algunos animales, poríferos y celentéreos, mantienen esta etapa, siendo animales diblásticos (con dos hojas blastodérmicas). Por ejemplo los pólipos tienen dos capas y se pueden asemejar a una gástrula invertida, siendo la mesoglea equivalente al blastocele y la cavidad interna en contacto con el exterior equivalente al arquénteron, no así el gastroporo con el blastoporo, pues tienen orígenes embrionarios diferentes. Estos animales son representantes de un nivel de organización muy sencillo que no han formado órganos sino algo parecido a tejidos.
Para que se hayan formado órganos se ha tenido que desarrollar una tercera hoja blastodérmica, aunque de tal forma que no se aumente demasiado el volumen, siguiendo la línea anteriormente descrita.
En la gástrula triblástica, células del endodermo se invaginan formando unas bolsas que se van ampliando hasta la consecución de una tercera hoja blastodérmica o mesodermo, incluida entre el endodermo y el ectodermo, con dos capas, una somatopleura cercana al ectodermo y otra esplacnopleura cercana al endodermo.
El mesodermo delimita junto con el mediastino (que dará lugar al mesenterio) una cavidad o celoma. Los animales con tres hojas blastodérmicas se denominan triblásticos, tanto acelomados, pseudocelomados como eucelomados. La formación del mesodermo por el proceso antes descrito se denomina enterocelia, pero no es el único mecanismo para va formación del mismo (véase mesodermo).